1 — миокардиоциты; 2 — анастомозы; 3 — вставочные участки; 4 — ядра миокардиоцитов; 5 — ядра эндотелия капилляров.
При электронной микроскопии в области вставочных дисков границы соседних клеток неровные: одна клетка вдается в другую пальцевидными выступами, что обеспечивает достаточную площадь сцепления клеток. По длине вставочного диска имеются различные по строению участки. Здесь много мест, содержащих волокнистое электроноплотное вещество, в которое вплетены концы тонких актиновых микрофиламентов (рис. 199). В зонах, не занятых микрофиламентами, расположены десмосомы и щелевые контакты. Считают, что в щелевых контактах происходит быстрая передача волны возбуждения от клетки к клетке без участия медиатора. Каждый сердечный миоцит содержит 1—2 ядра, расположенные в центре клетки, миофибриллы занимают периферическую часть цитоплазмы. Между миофибриллами одиночно, группами или цепочками расположены митохондрии, для которых характерно большое количество крист. Миофибриллы окружены системой трубочек и канальцев саркоплазматической сети. Развита Т-система, образованная трубчатыми впячиваниями плазмолеммы миоцита, причем Т-трубочки расположены на уровне Z-полоcок миофибрилл. В околоядерной зоне capкоплазмы расположены скопления митохондрий, комплекс Гольджи, лизосомы, гранулы гликогена и пигмента липофусцина.
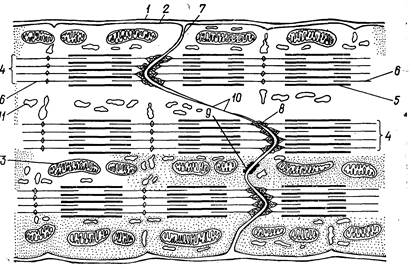
Рис. 199. Схема ультраструктурной организации миокардиоцитов в области контакта двух клеток (вставочного участка):
1 — сарколемма; 2 — плазмолемма; 3 — митохондрии; 4 — миофибрилла; 5 — миозиновые филаменты; б — актиновые филаменты; 7 — граница между миокардиоцитами; 8 — зона вплетения актиновых миофиламентов; 9 — десмосома; 10 — щелевой контакт; 11 — канальцы саркоплазматической сети.
Миоциты окружены тонковолокнистой соединительной тканью (эндомизий миокарда), в которой содержатся многочисленные капилляры, обеспечивающие процессы микроциркуляции. В сердце имеется опорный скелет, состоящий из фиброзных колец на границе между предсердиями и желудочками и в устьях сосудов, выходящих из желудочков. В составе фиброзных колец находятся пучки плотной коллагеновой соединительной ткани, а в сердце животных хрящевая и даже костная ткань (крупный рогатый скот). Фиброзно-хрящевые кольца препятствуют растяжению отверстий и обеспечивают прикрепление свободных концов волокон миокарда.
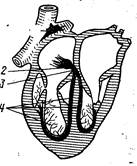 Автоматизм сокращения сердца, закономерная последовательность сокращений предсердий и желудочков на протяжении сердечного цикла обусловлены деятельностью атипичной мышечной ткани, входящей в состав проводящей системы сердца. Эта система состоит из синусно-предсердного узла, расположенного в устье краниальной полой вены, предсердно-желудочкового узла, лежащего в межпредсердной перегородке вблизи места прикрепления створки трехстворчатого клапана, предсердно-желудочкового ствола (пучка Гиса) и его разветвлений, расположенных под эндокардом межжелудочковой перегородки и в соединительнотканных прослойках миокарда — волокна Пуркине (рис. 200). Все эти компоненты проводящей системы образованы атипичными мышечными клетками, которые в функциональном отношении специализированы или на выработке импульса распространяющегося по всему сердцу и вызывающего сокращение его отделов в необходимой последовательности и с определенной частотой (клетки узлов), или на его проведении и передаче сократительным миоцитам.
Автоматизм сокращения сердца, закономерная последовательность сокращений предсердий и желудочков на протяжении сердечного цикла обусловлены деятельностью атипичной мышечной ткани, входящей в состав проводящей системы сердца. Эта система состоит из синусно-предсердного узла, расположенного в устье краниальной полой вены, предсердно-желудочкового узла, лежащего в межпредсердной перегородке вблизи места прикрепления створки трехстворчатого клапана, предсердно-желудочкового ствола (пучка Гиса) и его разветвлений, расположенных под эндокардом межжелудочковой перегородки и в соединительнотканных прослойках миокарда — волокна Пуркине (рис. 200). Все эти компоненты проводящей системы образованы атипичными мышечными клетками, которые в функциональном отношении специализированы или на выработке импульса распространяющегося по всему сердцу и вызывающего сокращение его отделов в необходимой последовательности и с определенной частотой (клетки узлов), или на его проведении и передаче сократительным миоцитам.
Рис. 200. Схема проводящей системы сердца:
1 — синусно-предсердный узел; 2 — предсердно-желудочковый узел; 3 — предсердно-желудочковый ствол (пучок Гиса); 4 — его ножки и разветвления.
Атипичные миоциты проводящей системы имеют характерные микроскопические и ультраструктурные признаки, отличающие их от сократительных миоцитов. При обычной гематоксилиновой окраске они более светлые, имеют неправильно овальную форму и, как правило, поперечный диаметр их больше, чем диаметр сократительных миоцитов, в 2—3 раза. Однако в составе синусно-предсердного узла обнаружены мелкие клетки округлой формы. В функциональном отношении это водители ритма — пейсмекеры. Весьма характерным для атипичных миоцитов являются большой объем саркоплазмы и слабое развитие миофибриллярного аппарата. Миофибриллы занимают самую периферическую часть в цитоплазме клеток, не имеют параллельной ориентации, вследствие чего для атипичных миоцитов несвойственна поперечная исчерченность. У них слабо развит саркоплазматический ретикулум, отсутствует система Т-трубок, а в саркоплазме мало митохондрий, но имеется большое количество гранул гликогена. В этих клетках много гликолитических ферментов и уменьшенное количество ферментов аэробного окисления (сукцинатдегидрогеназы и цитохромоксидазы), что свидетельствует о преобладании в них анаэробного гликолиза. Клетки проводящей системы значительно более устойчивы к кислородному голоданию, чем сократительные миоциты.
Перейти на страницу:
1 2 3