Средняя часть глазного бокала. При проколе бокала регистрировали небольшой положительный ПП до двух мВ, который при длительном нахождении электрода в этой точке мог падать до нуля. ЭРГ, полученные в этих точках, имели вид волн, среди которых встречались как положительные, так и отрицательные. Были зарегистрированы двух- и трехфазные волны ЭРГ (рис. 1б 1 – 3) При дальнейшем погружении микроэлектрода ПП падал до нулевого значения, вместе с тем уменьшалась и амплитуда ЭРГ без заметного искажения фаз. В этой части бокала было получено 69 ЭРГ.
Нейропиль. На первых этапах погружения электрода в глазной бокал наблюдался отрицательный ПП величиной до 10 мВ. В этой точке регистрировали ЭРГ, по крайней мере, трех видов.
1. Отрицательная монофазная волна амплитудой до 7 мВ, состоящая из относительно быстрого смещения потенциала и более медленного его возврата к исходному значению (реполяризация) (рис. 2 4). Такой вид имели около 45 % из 99 зарегистрированных в этой точке ЭРГ.
2. В стольких же случаях регистрировали сходную по кинетике положительную волну ЭРГ амплитудой до 8 мВ (рис. 2 3).
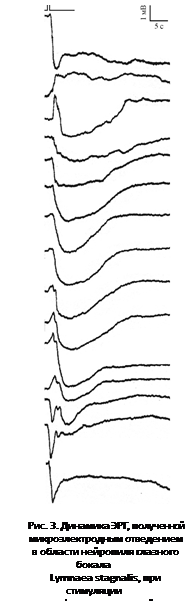
3. В 10 % случаев медленные волны ЭРГ сочетались с быстрыми колебаниями, не привязанными к определенной фазе волны. Среди них быстрые отрицательные (рис. 2 6) и положительные пики (рис. 2 2, 5) с отрицательной медленной последующей волной, а также множества положительных пиков на фоне общей медленной деполяризации (рис. 2 1). С течением времени форма ЭРГ этих пиков могла изменяться. Наблюдаемые изменения касались как амплитуды, так и (иногда) полярности компонентов ответов (рис. 3). При дальнейшем погружении электрода сначала наблюдали увеличение ПП и амплитуды фаз ЭРГ, а затем – уменьшение обоих сигналов.
Обсуждение
 Задача регистрации электрических сигналов от различных участков изолированного глаза брюхоногого моллюска ставилась и прежде. Исследованиями на Strombus luhuanus [6] и Helix aspersa [7] показано, что полярности фаз ЭРГ, получаемых электродами-присосками от основания глазного бокала и области роговицы, противоположны. Перемещение электрода внутри глазного бокала от микровиллярного слоя глаза H. aspersa к роговице также сопровождается инверсией полярности волны ЭРГ с отрицательной на положительную [7]. Изменение полярности фаз ЭРГ в зависимости от положения регистрирующего электрода-присоски наблюдалась также и в исследованиях на Achatina fulica. При этом в области боковой поверхности бокала была получена двухфазная ЭРГ, состоящая из быстрого пика положительной и последующей медленной отрицательной волны [8]. В своих опытах мы получили результат, противоречащий на первый взгляд приведенным данным. Так, в независимых экспериментах при погружении микроэлектрода в топографически близкие точки глазного бокала могла быть получена различная полярность компонентов ЭРГ. Более того, как оказалось, при длительной регистрации в одном и том же положении микроэлектрода форма и полярность компонентов ЭРГ могли претерпевать изменения. Причину наблюдаемой динамики можно усматривать в том, что в разных отведениях кончик микроэлектрода был погружен в сетчатку на различную глубину. При продолжительной же регистрации силы деформации мягкой ткани глаза, возникшие при погружении микроэлектрода, могли вызывать ее постепенное движение и, таким образом, смещение глубины точки регистрации относительно первоначального положения в бокале. Если такое рассуждение справедливо, то следует признать наличие зависимости амплитуды и полярности ЭРГ от глубины погружения электрода, как это было показано на примере глаза насекомых, который по сравнению с моллюсками имеет более жесткую структуру, дающую возможность послойного анализа электрических сигналов. Таким методом был получен профиль постоянного потенциала, коррелирующий с амплитудой ЭРГ [9]. Для объяснения существования пиков в профиле постоянного потенциала, а также точки реверсии амплитуды ЭРГ был использован подход к глазу насекомого как объемному проводнику, содержащему много источников тока. Предполагалось существование нескольких контуров темнового тока, разбалансировка которых при освещении приводит к возникновению компонентов ЭРГ. В частности, такие токи могут генерироваться последовательно расположенными на пути погружения микроэлектрода фоторецепторами и нейронами ламины. В структуре сетчатки L. stagnalis можно указать, по крайней мере, на два типа клеток, которые могли бы генерировать токи и вносить вклад в формирование ЭРГ: непосредственно фоторецепторные и нервные клетки [2]. Ультраструктурными исследованиями функциональная связь между этими клетками до сих пор не установлена [2, 10], и предположение о том, что в сетчатке L. stagnalis именно в клетках второго порядка формируется импульсный сигнал в ответ на освещение сетчатки, основывается на электрофизиологических данных [11]. Сам вопрос о том, в каких клетках глаза брюхоногих моллюсков происходит генерация распространяющегося потенциала действия, является важным для понимания функциональной организации сетчатки этих животных. Импульсная активность фоторецепторных клеток обнаружена в сетчатке голожаберных моллюсков Aplysia californica [12] и Hermissenda crassicornis [13, 14], а также в светочувствительных клетках малого ганглия заднежаберного Clione limacina [15]. Фоторецепторы S. luhuanus [16] и A. fulica [8] отвечают на освещение лишь градуальными изменениями мембранного потенциала и импульсный ответ формируется вероятнее всего в клетках второго порядка. Нам не удалось обнаружить явных признаков потенциалов действия в электрических ответах на освещение глаза L. stagnalis, хотя в составе ЭРГ, полученной от области нейропиля, часто присутствовали быстрые пики, появление которых может быть обязано синхронному возбуждению нейронов. Можно полагать, что зоны генерации потенциалов действия локализованы главным образом как раз в этой области. Такое предположение может объяснить тот факт, что ЭРГ, полученные в области роговицы или средней части глазного бокала, практически не содержат быстрых пиков. Во всяком случае сравнительно независимое изменение быстрых и медленных компонент ЭРГ наводит на мысль о том, что они могут генерироваться различными элементами сетчатки.
Задача регистрации электрических сигналов от различных участков изолированного глаза брюхоногого моллюска ставилась и прежде. Исследованиями на Strombus luhuanus [6] и Helix aspersa [7] показано, что полярности фаз ЭРГ, получаемых электродами-присосками от основания глазного бокала и области роговицы, противоположны. Перемещение электрода внутри глазного бокала от микровиллярного слоя глаза H. aspersa к роговице также сопровождается инверсией полярности волны ЭРГ с отрицательной на положительную [7]. Изменение полярности фаз ЭРГ в зависимости от положения регистрирующего электрода-присоски наблюдалась также и в исследованиях на Achatina fulica. При этом в области боковой поверхности бокала была получена двухфазная ЭРГ, состоящая из быстрого пика положительной и последующей медленной отрицательной волны [8]. В своих опытах мы получили результат, противоречащий на первый взгляд приведенным данным. Так, в независимых экспериментах при погружении микроэлектрода в топографически близкие точки глазного бокала могла быть получена различная полярность компонентов ЭРГ. Более того, как оказалось, при длительной регистрации в одном и том же положении микроэлектрода форма и полярность компонентов ЭРГ могли претерпевать изменения. Причину наблюдаемой динамики можно усматривать в том, что в разных отведениях кончик микроэлектрода был погружен в сетчатку на различную глубину. При продолжительной же регистрации силы деформации мягкой ткани глаза, возникшие при погружении микроэлектрода, могли вызывать ее постепенное движение и, таким образом, смещение глубины точки регистрации относительно первоначального положения в бокале. Если такое рассуждение справедливо, то следует признать наличие зависимости амплитуды и полярности ЭРГ от глубины погружения электрода, как это было показано на примере глаза насекомых, который по сравнению с моллюсками имеет более жесткую структуру, дающую возможность послойного анализа электрических сигналов. Таким методом был получен профиль постоянного потенциала, коррелирующий с амплитудой ЭРГ [9]. Для объяснения существования пиков в профиле постоянного потенциала, а также точки реверсии амплитуды ЭРГ был использован подход к глазу насекомого как объемному проводнику, содержащему много источников тока. Предполагалось существование нескольких контуров темнового тока, разбалансировка которых при освещении приводит к возникновению компонентов ЭРГ. В частности, такие токи могут генерироваться последовательно расположенными на пути погружения микроэлектрода фоторецепторами и нейронами ламины. В структуре сетчатки L. stagnalis можно указать, по крайней мере, на два типа клеток, которые могли бы генерировать токи и вносить вклад в формирование ЭРГ: непосредственно фоторецепторные и нервные клетки [2]. Ультраструктурными исследованиями функциональная связь между этими клетками до сих пор не установлена [2, 10], и предположение о том, что в сетчатке L. stagnalis именно в клетках второго порядка формируется импульсный сигнал в ответ на освещение сетчатки, основывается на электрофизиологических данных [11]. Сам вопрос о том, в каких клетках глаза брюхоногих моллюсков происходит генерация распространяющегося потенциала действия, является важным для понимания функциональной организации сетчатки этих животных. Импульсная активность фоторецепторных клеток обнаружена в сетчатке голожаберных моллюсков Aplysia californica [12] и Hermissenda crassicornis [13, 14], а также в светочувствительных клетках малого ганглия заднежаберного Clione limacina [15]. Фоторецепторы S. luhuanus [16] и A. fulica [8] отвечают на освещение лишь градуальными изменениями мембранного потенциала и импульсный ответ формируется вероятнее всего в клетках второго порядка. Нам не удалось обнаружить явных признаков потенциалов действия в электрических ответах на освещение глаза L. stagnalis, хотя в составе ЭРГ, полученной от области нейропиля, часто присутствовали быстрые пики, появление которых может быть обязано синхронному возбуждению нейронов. Можно полагать, что зоны генерации потенциалов действия локализованы главным образом как раз в этой области. Такое предположение может объяснить тот факт, что ЭРГ, полученные в области роговицы или средней части глазного бокала, практически не содержат быстрых пиков. Во всяком случае сравнительно независимое изменение быстрых и медленных компонент ЭРГ наводит на мысль о том, что они могут генерироваться различными элементами сетчатки.
Перейти на страницу:
1 2 3 4