Опыты выполнены на белых беспородных крысах обоего пола весом 120-200 г, оперированных под легким эфирным наркозом, а затем обездвиженных тубокурарином (1,5 мг/кг в/м), и переведенных на искусственное дыхание. Трахеотомия не производилась, воздух в легкие нагнетался через ноздри. Операционное поле подвергалось местной анестезии 0,5% раствором новокаина. Осуществлялась жесткая фиксация головы с помощью головодержателей. Череп трепанировался над ПМСБ корковой области С1 по координатам: 2-3 мм каудальнее брегмы и 5-6 мм латеральнее сагиттального шва. Импульсная активность корковых нейронов, а также фокальные ВП отводились стеклянными одиночными микроэлектродами, заполненными 2,5 М раствором NaCL, с сопротивлением около 5 мОм и вольфрамовыми в стеклянной изоляции полумикроэлектродами с сопротивлением 0,5-1 мОм, которые погружались в кору отвесно к ее поверхности. Контролем попадания электродов в ПМСБ служили фокальные ВП, предварительно прослушиваемые на аудиомониторе, которые возникали в ответ на легкое постукивание по вибриссам контралатеральной стороны мордочки крысы вначале вручную, а затем с помощью специальных тактильных датчиков.
Идентификация колонок. Сравнивая фокальные ВП, наблюдаемые на видеомониторе по их латентным периодам (ЛП), крутизне и амплитуде в ответ на сгибание вибрисс, устанавливали, как показано на рис.1,а, соответствие между бочонковой колонкой и центральной в РП. Для более точной идентификации центра РП нейронов производилась стимуляция ближайших к центру вибрисс со ступенчатым уменьшением амплитуды их отклонения (рис.1,б). Наблюдаемое при этом сужение РП до одной вибриссы в случае ее центрального положения имело место при наиболее низких амплитудах ее отклонения.
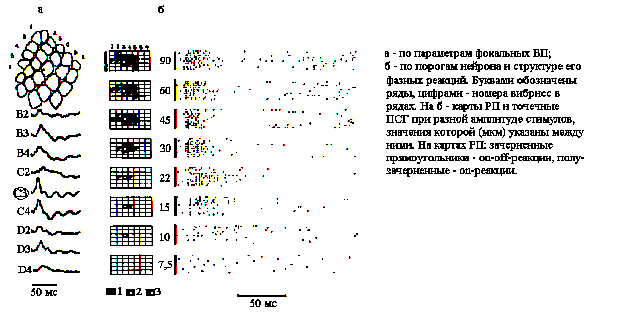
Рис. 1. Идентификация бочонковых колонок С1 коры мозга крысы.
Картирование РП.
Картирование РП нейронов производилось путем построения постстимульных гистограмм (ПСГ) реакций нейронов на последовательное отклонение вибрисс вначале контралатеральной, а затем ипсилатеральной стороны мордочки крысы. Амплитуда отклонения вибрисс при картировании превышала в десять раз ее пороговое значение, определяемое для центра РП.
Определение порога. При определении пороговых характеристик нейронов в качестве адекватного стимула использовано сгибание центральной в РП вибриссы с удержанием ее в отклоненном положении в течение 0,1-0,5 с. Чтобы избежать неконтролируемых смещений, более дальние от носа вибриссы, достигающие в длину 30-50 мм, укорачивались справа и слева до 5-10 мм. Механический датчик для определения пороговых характеристик нейронов представляет собой щуп, приклеенный на свободном конце пьезокерамической пластины, изготовленной в ОКБ "Пьезоприбор" РГУ, на которую подавалось напряжение от 60 до 1В с выхода электростимулятора ЭСЛ-2, задающего амплитуду отклонения на самом дальнем конце щупа в диапазоне от 90 до 1,5 мкм. Предварительная метрологическая проверка показала линейный характер зависимости между напряжением и амплитудой сгибания пьезокерамической пластины в используемом диапазоне. Поэтому непосредственно в опыте амплитуда отклонения щупа оценивалась по напряжению на выходе ЭСЛ-2. Серия стимулов определенной амплитуды предъявлялась ритмично с частотой 0,5 Гц. По 10-20 реализациям строились точечные и столбиковые ПСГ с бином 1-2 мс за период 100 или 500 мс. При определении пороговых характеристик нейронов рассматривались только on-эффекты стимуляции. Определялись параметры первичных компонентов возбудительных (В-) реакций на каждый из стимулов серии, а также усредненный паттерн разряда. Пороговым считалось такое значение амплитуды, при котором нейрон реагировал в серии повторных стимулов с вероятностью равной или большей 0,5.