Существует несколько гипотез о механизмах интерференции, но ни одна из них не является общепринятой и ни одна из них мне не нравится. Поэтому я лучше приведу предложенную И.П.Горловым [8] гипотезу о том, зачем нужна интерференция. Суть ее такова. Ничего в природе не бывает бесплатно. Конечно, рекомбинация дает определенные преимущества, но она обходится и организму недешево. Чего стоит одно разрезание живой ДНК! Естественный отбор должен балансировать рекомбинацию таким образом, чтобы ее преимущества превышали затраты. Два обмена стоят дороже, чем один, где бы они ни были расположены. Но если мы подумаем о преимуществах, то станет понятно, что чем ближе они друг к другу, тем менее полезны в смысле перетасовки генов. Второй обмен в непосредственной близости от первого практически аннулирует его эффект, возвращая гомологичные хромосомы в исходное состояние (рис.9). Поэтому, если уж тратиться на обмены, то надо расположить их подальше друг от друга. Что и происходит благодаря интерференции, только до сих пор непонятно как.
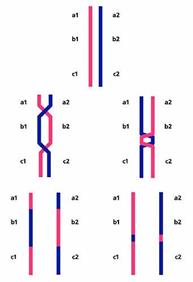
Рис.9. Результаты двух рекомбинационных событий на далеком (слева) и близком (справа) расстоянии друг от друга. В первом случае рекомбинантные хромосомы отличаются от родительских по набору генов a, b, c; во втором - не отличаются.
Все перечисленные выше правила так или иначе обусловлены позиционными эффектами, которые играют, видимо, главную роль в распределении обменов по хромосомам.
Правило светлого района связано с особенностями упаковки ДНК мейотических хромосом. Отдельные районы хромосом сильно отличаются по этому признаку. Хромосомы млекопитающих после обработки красителями, специфично связывающимися с ДНК, выглядят поперечно-полосатыми. В интенсивно окрашенных (темных) участках концентрация ДНК выше, чем в светлых. Особенности упаковки зависят от генетического состава ДНК. Сильно огрубляя ситуацию, можно сказать, что гены сосредоточены в основном в светлых районах, а темные главным образом состоят из бессмысленных и сильно повторенных последовательностей ДНК. Но это очень грубая картина. Гены есть и в темных районах, а в светлых довольно много бессмысленных последовательностей.
Правило светлого района состоит в том, что рекомбинация происходит в таких районах чаще, чем в темных. Однако пока не ясно почему. Можно думать, что ДНК светлых районов более активно участвует в поиске гомологии, в ней чаще возникают опознавательные связки и рекомбинационные обмены. На это указывает тот факт, что в мейотических хромосомах относительный размер светлых районов непропорционально больше, чем в митотических, а темных - непропорционально меньше. До наших экспериментов этот факт базировался на косвенных данных*. Было известно, что хромосомы, содержащие много светлых районов, в мейозе оказываются относительно более длинными, чем в митозе. Нам впервые удалось это увидеть, применив дифференциальное окрашивание мейотических хромосом обыкновенной бурозубки. Светлые районы были длиннее.
Правило светлого района на первый взгляд кажется нецелесообразным. Цель рекомбинации состоит в перетасовке генов. Но из этого не следует, что рекомбинация должна происходить именно в тех районах, где сконцентрированы гены. Гораздо логичней было бы производить обмены там, где генов нет. Результат был бы тем же, а риск повреждения генов за счет их разрезания, спаривания свободных цепей, подчистки результатов неверного спаривания был бы сведен к нулю. Однако логика эволюции не всегда совпадает с логикой инженера. Она (эволюция) никогда не создает механизмы с чистого листа. Она предпочитает слегка модифицировать уже существующие.
Мы уже говорили, что большинство белков, обеспечивающих рекомбинацию, - это гомологи бактериальных и эукариотических белков, участвующих в репарации, залечивании мутационных повреждений ДНК. Поскольку рекомбинация возникла в эволюции позже репарации и использует слегка измененную машину репарации, можно заключить, что она и произошла от репарации. Более того, скорее всего на первых этапах, когда еще не было мейоза, она была всего лишь одним из вариантов репарации, предназначенным для залечивания самых опасных и тяжелых повреждений ДНК - двунитевых разрывов. Если разорвана одна нить, разрыв можно залечить, используя вторую нить в качестве матрицы. Если же разорваны две нити, нужно найти гомологичный участок ДНК в другом месте генома, расплести его и использовать как матрицу (рис.4).
С этой точки зрения становится понятной и нецелесообразная на первый взгляд концентрация обменов в светлых, богатых генами районах хромосом, и кажущаяся избыточность разрывов ДНК на ранних стадиях точного опознавания гомологов, и тот факт, что только малая часть этих разрывов превращается в обмены, а большая - ни к каким обменам не ведет. В ходе подчистки необменных связок может происходить исправление потенциальных повреждений в генах.
Перейти на страницу:
1 2 3