Далее рост боковой ветви осуществляется так же, как и ствола, т. е. удлинение ее происходит непосредственно под гидрантом, а сам гидрант постепенно увеличивается в размерах. Со временем и боковая ветвь также начинает ветвиться. Определенной ориентации ветвления нет, хотя иногда ветви отходят преимущественно от одной стороны ствола, а иногда поочередно от противоположных. Возможны и другие варианты.
Перманентный макияж
Первоначально боковая ветвь перпендикулярна к той, от которой она отходит, но затем она загибается вверх. Эксперименты с регенерировавшими кусками ствола показали, что переворачивание такой модельной колонии гидрантом вниз приводит по мере дальнейшего роста к изгибанию ствола вместе с венчающим его гидрантом вверх. Вследствие этой особенности большинство ветвей колонии оказывается ориентированной более-менее параллельно друг другу (рисунок, 3).
Скорости роста всех ветвей и ствола примерно одинаковы и составляли около 0,5 мм/сут при температуре 12°, что соответствует данным Маки (Mackie, 1966) для Т. сгосеа при 14°—1 мм/сут. Из-за сходства скоростей роста и определенного порядка образования ветвей на некотором расстоянии от апикально расположенного гидранта длина ветвей пропорционально возрастает в проксимальном направлении, а венчающие их гидранты оказываются примерно на одном уровне. Эта особенность морфологии и роста колонии у Т. larynx объясняет, почему в кусте гидранты расположены преимущественно на его поверхности или вблизи ее (рисунок, 7).
Гидрориза у Т. larynx морфологически слабо отличается от побегов. Столоны лишь некоторое время стелются по субстрату, а затем теряют с ним связь и превращаются в стволы побегов. При этом на их верхушках роста формируются терминальные гидранты. Благодаря тому, что столоны ветвятся чаще, чем стволы и ветви побегов, в среднем через каждые 2 мм, связь колонии с субстратом обеспечивается достаточно прочная, несмотря на то, что площадь, занятая гидроризой, относительно мала. По данным Берилла (Berrill, 1952). у Т. сгосеа гидрориза клубковидная. В зрелых колониях верхушки роста столонов встречаются редко.
Кроме функции прикрепления к субстрату гидрориза у Т. larynx выполняет и другую важную функцию—хранение клеточного материала в период переживания колонией неблагоприятных условий. На Белом море всю долгую зиму с октября по июнь колонии Т. larynx существуют в форме столонов и остатков стволов, причем в первых сосредоточен основной клеточный материал, за счет которого и начинается рост колонии летом следующего года.
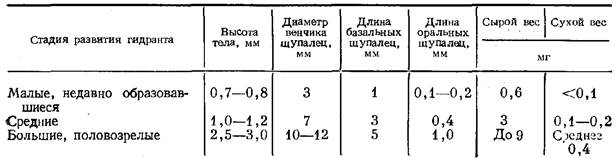
Как известно (Тихомиров, 1887; Наумов, 1960; Allman, 1871; Hyman 1940), гидранты рода Tubularia обладают двумя венчиками щупалец — оральным и аборальным (или базальным), причем последний расположен в нижней трети гидранта и обладает значительно более длинными щупальцами, чем оральный венчик. Венчик базальных щупалец имеет в основании хорошую опору в виде кольца энтодермальной паренхимы (Тихомиров, 1887). Нетрудно увидеть в этом приспособление для обитания на сильном течении. Кроме того, при определенной концентрации гидрантов, расположенных на поверхности куста, они поддерживают друг друга.
Мы видим, что значительные размеры гидрантов, наличие у них двух венчиков щупалец, определенная закономерность ветвления и роста ветвей, обеспечивающая расположение гидрантов примерно на одном уровне, своеобразный ограниченный рост гидроризы, способствующий концентрации колонии на одном месте,— все это оказывается функционально связанным друг с другом, обеспечивающим в совокупности приспособление этого вида к обитанию на течении. Лишь немногие виды гидроидов способны вместе с Т. larynx противостоять течению в желобе Великой Салмы; это—Hydrallmania falcata (L.), Abietinaria abietina (L.), Sertularia mirabilis (Verrill).