Размер и форма РП нейронов бочонковых колонок
Картированы В-зоны РП 82 нейронов разных уровней бочонковых колонок ПМСБ С1 коры мозга крыс. Исследования проводили в плане сравнения размеров и формы РП нейронов, находящихся в представительстве латеральных (преимущественно в бочонке С3) и медиальных (бочонки рядов В и А) вибрисс. В первой выборке нейронов (N=52), представляющей бочонок С3, размер РП при амплитуде стимула, равной десяти пороговым значениям, как это показано на рис.2а, варьировал от одной до 23 вибрисс. Около 11% нейронов, относящихся, преимущественно, к слою 4, имели одновибриссные РП. Амплитудные пороги у этих нейронов были наиболее низкими - от 3 до 15 мкм, при среднем значении 5,3+0,7 мкм.
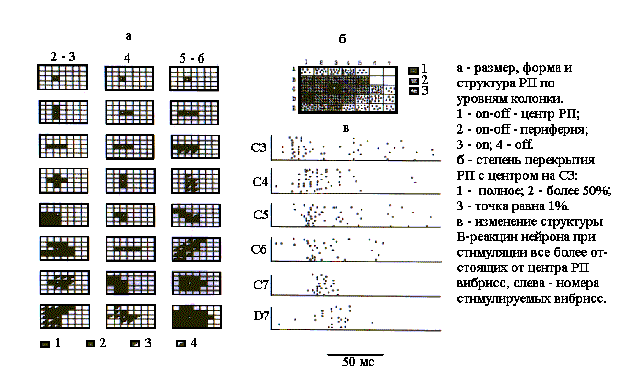
Рис. 2. РП нейронов разных уровней бочонковой колонки С1.
Нейроны поверхностных (2-3) и глубоких (5-6) слоев имели многовибриссные РП, которые всегда были сплошными. У 23% нейронов РП имели вытянутую, чаще всего вдоль, но иногда и поперек вибриссковых рядов, форму. В последнем случае угол наклона РП к рядам вибрисс варьировал у разных нейронов, но чаще всего был прямым. Однородную структуру РП с on-off-ответами имели 31%, а с on-ответами - 8% нейронов. Большинство нейронов имели неоднородные РП: с on-off-ответами в центре и с оn-ответами по краю - 23%, а с on-off-центром и off-краем - 38% нейронов. При этом у 70% нейронов центр РП располагался эксцентрически, т.е. находился вблизи края В-зоны РП. Средние значения амплитудных порогов у нейронов с многовибриссными РП были достоверно (Р<0,01) выше, чем у одновибриссных: на уровне поверхностных слоев - 9,4+0,3 мкм, а в глубоких слоях - 13,0+0,8 мкм. Нейроны, РП которых перекрывали почти все контралатеральное поле вибрисс, имели амплитудные пороги около 30 мкм.
РП нейронов, принадлежащих одной и той же бочонковой колонке, существенно перекрывались. Центры РП данной выборки нейронов всегда совпадали, поскольку это было исходной предпосылкой для отнесения регистрируемых нейронов к бочонковой колонке С3. Наложением картированных РП произведена количественная оценка степени их перекрытия во всей выборке нейронов, представляющих колонку С3 (рис.2,б). Анализ показал, что все вибриссы контралатеральной стороны в той или иной мере адресованы нейронам данной колонки. При этом степень перекрытия РП градуально уменьшилась к краям вибрисскового поля до 5%. Однако, подзона с более чем 50-процентным перекрытием РП, заштрихованная на рис.2б, имела симметричную и вытянутую вдоль ряда С форму. Таким образом, суммарное РП нейрональной колонки С3 ориентировано вдоль одноименного ряда вибрисс, в плоскости которого совершаются их ощупывающие движения, и имеет значительное перекрытие с РП нейронов соседних бочонковых колонок этого, а также верхнего и нижнего рядов.
На рис.2,в в качестве примера более общей закономерности показано удлинение ЛП, уменьшение частоты первичного разряда, а также перестройка структуры В-реакции нейрона, зарегистрированного на уровне слоя 4, при поочередной стимуляции вибрисс, отстоящих на все большем расстоянии от центра РП. В среднем по выборке (N=52) ЛП при смещении стимула от центра на периферию уменьшался с 9,0+0,3 мс до 19,1+1,2 мс. Уменьшалась при этом также и частота первичного разряда в первые 50 мс после отметки стимула с 132+21 имп/с до 41+8 имп/с. Все это в совокупности свидетельствует об уменьшении функциональной эффективности синаптических влияний на колонку по направлению к периферии РП ее нейронов.
На рис.1,б показана структура В-зоны РП нейрона, зарегистрированного на глубине 1100 мкм и принадлежащего слою 5, при разных амплитудах тактильного стимула, а также соответствующие им точечные ПСГ (отклонялась вибрисса С3). При 10-пороговой амплитуде (90 мкм) В-зона РП данного нейрона состояла из 14 вибрисс, 6 из которых вызывали on-off-ответы и находились в центре РП, а 8 периферических вибрисс - on-ответы. При уменьшении амплитуды количество вибрисс в В-зоне РП уменьшалось вначале незначительно и, в основном, за счет периферии. Значительное уменьшение площади РП наблюдалось у этого и других нейронов при амплитудах 30 и 22 мкм и совпадало с исчезновением в структуре В-реакции нейрона на отклонение даже центральной вибриссы компонентов вторичной активации.
Перейти на страницу:
1 2 3 4 5 6