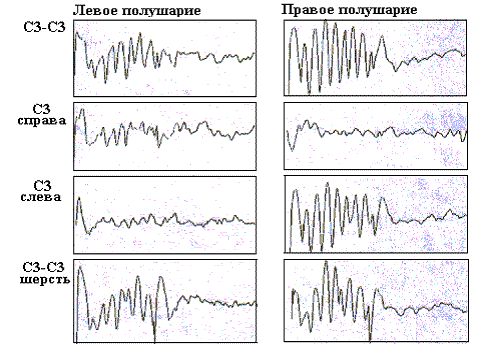
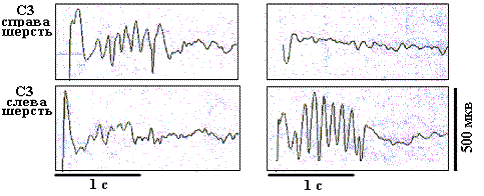
Рис. 8. Усредненные ВП БК бочонковых колонок С1 коры мозга крыс, регистрируемые на уровне афферентных слоев, при разном положении билатеральных и унилатеральных тактильных стимулов.
Есть основания, в связи с этим, полагать, что вторичные разряды нейронов в фокальной бочонковой колонке синхронизованы по ансамблю четче, чем на периферии проекционного поля. Следовательно, ассоциативные взаимодействия, проявляющиеся в ритмических разрядах, назначение которых состоит в интегрировании специфической информации о пространственных и временных событиях в РП нейронов поля бочонков, осуществляются, главным образом, через фокальные проекционные модули.
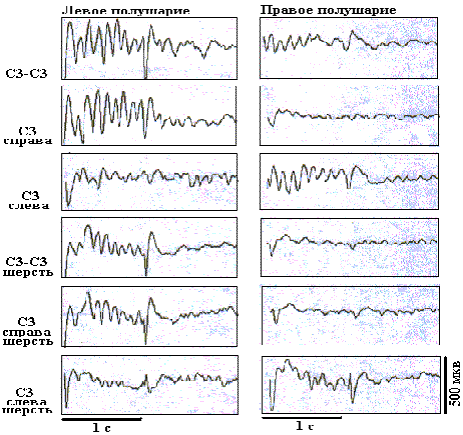
Рис. 9. Усредненные ВП БК бочонковых колонок С1 коры мозга крыс, регистрируемые на уровне эфферентных слоев, при разном положении билатеральных и унилатеральных тактильных стимулов.
На рисунках 8 и 9 представлены усредненные фокальные ВП в БК колонок С3 при разном пространственном положении тактильных стимулов, предъявляемых унилатерально и билатерально. Использовалось изолированное 10-пороговое отклонение вибрисс, а также отклонение вибрисс у их основания, при котором стимулировалась также и шерсть. Для уровня афферентных слоев отмечается четкая индивидуальность паттернов фокальных ВП в БК колонок, соответствующая каждому из условий стимулирования. Так, при билатеральном изолированном отклонении вибрисс в бочонковых колонках С3 правого и левого полушарий наблюдаются коротколатентные и высокоамплитудные негативные первичные волны и до семи вторичных ритмических негативно-позитивных колебаний. При этом колонка в правом полушарии доминирует по общему уровню возбудимости, что проявляется в более высокой амплитуде вторичных волн. Это доминирование имело место на уровне афферентных слоев при разном пространственном положении тактильных стимулов, когда они были адресованы правому полушарию.
Унилатеральная изолированная стимуляция вибрисс вызывала в БК латерализованные контралатеральные первичные и вторичные коротколатентные потенциалы. При билатеральной стимуляции шерсти возникали более короткие по латентности ВП в каждой из колонок БК. При унилатеральной стимуляции шерсти ВП возникали в обоих полушариях, но в ипсилатеральной колонке их латентность была в два раза большей (около 20 мс). На уровне эфферентных слоев паттерны ВП не имели такой четкой специфичности, как в афферентных слоях. На выходном уровне гомотопных колонок при всех положениях тактильных стимулов доминирующим было левое полушарие, что проявилось в более высокой, по сравнению с правым полушарием, амплитуде фокальных ВП левой колонки.
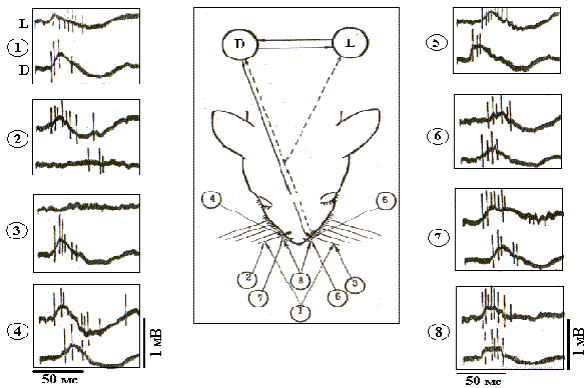
Рис. 10. Фокальные ВП и импульсные реакции нейронов БК бочонковых колонок при разном пространственном положении унилатеральных и билатеральных тактильных стимулов.
Индивидуальный профиль фокальных ВП и импульсных реакций нейронов БК бочонковых колонок при разном пространственном положении тактильных стимулов более наглядно представлен на рис. 10. На электрограммах показаны первичные компоненты В-реакций нейронов из симметричной пары колонок С3, и негативные первичные волны ВП при более широкой полосе пропускания фильтров на входе электроэнцефалографа (2-2000 Гц) - фокальные ВП. В левой (L) и правой (D) колонках наблюдаются по два нейрона, различающиеся по амплитуде регистрируемых спайков. Видно, что вызванная активность на стимуляцию шерсти характеризуется более короткой латентностью в контралатеральной колонке, а при унилатеральном изолированном отклонении вибрисс ответы наблюдаются только в контралатеральном полушарии.