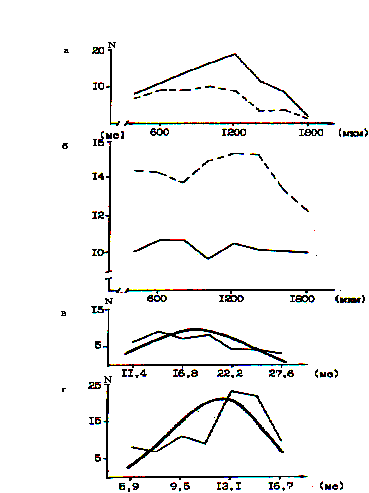
Рис.6. Послойное распределение в бочонковой колонке нейронов с билатеральными и контралатеральными афферентными входами, а также ЛП их В-реакций на стимуляцию центра РП (объяснения в тексте).
Тонкими линиями изображены эмпирические распределения ЛП реакций, соответственно, БВ- и КВ-нейронов. Плавные (толстые линии) графики получены в результате выравнивания эмпирических вариационных рядов по нормальному закону. Оценка различий между эмпирическими и теоретическими распределениями значений ЛП В-реакций по критерию Пирсона "хи-квадрат" показала, что для БВ-нейронов распределение ЛП является нормальным, а для КВ-нейронов отличается от нормального (Р<0,001). В связи с этим необходимо отметить, что среди КВ-нейронов выделяются две крайние группы: малочисленная с коротколатентным возбуждением и более многочисленная с длиннолатентным возбуждением. Между ними находится наиболее многочисленная группа нейронов, характеризующаяся прямолинейной регрессией между количеством возбужденных нейронов и значением латентности. Эти данные свидетельствуют о том, что, с одной стороны, в пространственно разнесенной по бочонковой колонке группе входных нейронов, имеется отчетливый градиент функциональной эффективности влияния первичных таламических афферентов, а, с другой стороны, имеет место последовательная полисинаптическая передача возбуждения по направлению к поверхностным и глубоким слоям коры мозга.
Входы от РП шерсти наиболее эффективно проявляют себя в глубоких и средних слоях и нормальный характер распределения ЛП реакций БВ-нейронов свидетельствует как о слабо выраженном градиенте их функциональной эффективности по колонке, так и о незначительном ветвлении цепочек БВ-нейронов.
Поскольку котралатеральный лемнисковый тракт в системе тригеминоталамических путей крысы полностью перекрещивается, а дорсальный ипсилатеральный тракт отсутствует /25/, была исследована возможность передачи ипсилатеральной афферентации через мозолистое тело. Тем более что на это указывали достоверно более высокие значения ЛП реакций БВ-нейронов на стимуляцию шерсти, свидетельствующие о непрямом ее поступлении в ПМСБ. В ряде опытов с этой целью регистрировались реакции нейронов бочонковых колонок на ипсилатеральную стимуляцию вибрисс и шерсти в контроле, а также после обратимой депрессии (с помощью аппликации раствора KCl) контралатеральной области С1, либо после перерезки мозолистого тела в проекции сагиттального шва. Во всех случаях, когда удавалось удержать нейрон при разобщении полушарий, характер реагирования нейронов на контралатеральные и ипсилатеральные стимулы сохранялся, что
указывает на наличие не опосредованного мозолистым телом пути ипсилатеральной афферентации к бочонковым колонкам представительства латеральных вибрисс в С1 коры мозга крыс. В ряде случаев наблюдалось достоверное (Р<0,01) увеличение ЛП нейронных реакций на ипсилатеральную стимуляцию, что указывает на облегчающий характер влияния через мозолистое тело на нейроны бочонковых колонок.
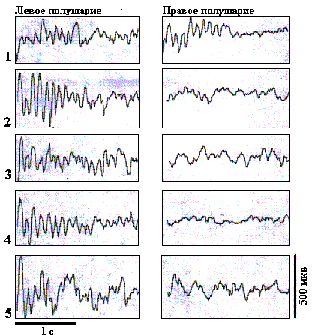
Рис. 7. Усредненные ВП БК бочонковых колонок С1 коры мозга крыс при разном положении тактильного стимула в правом поле вибрисс.
Отклонение вибрисс: 1- С2, 2- С3, 3- С4, 4- В3, 5- D3.
Фокальные ВП в БК бочонковых колонок при разном пространственном положении тактильных стимулов.
На рис.7 представлены усредненные по 40 реализациям фокальные ВП БК бочонковых колонок в представительстве латеральных вибрисс С3 слева и справа при разном унилатеральном положении тактильного стимула. В левом, контралатеральном по отношению к стимулу (10-пороговому отклонению вибриссы С3 справа), полушарии на уровне афферентных слоев бочонковой колонки С3 наблюдались разной выраженности и латентности ВП с вторичными ритмическими послеразрядами. ЛП и амплитуда негативной первичной волны ВП в ответ на стимуляцию центральной в РП вибриссы были наиболее оптимальными. Но более характерна для центрального положения стимула четкая структура ритмических послеразрядов на включение наличие ВП на выключение. Это коррелирует с более высокой структурированностью вторичной активации в фазных нейронных реакциях на стимуляцию центра РП, которая проиллюстрирована на рис.5.