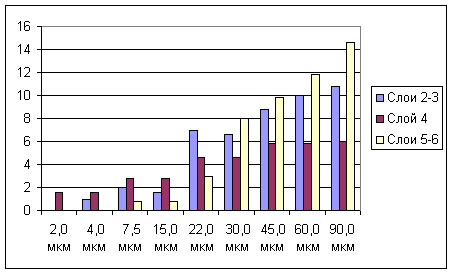
Рис. 3. Зависимость размеров В-зоны РП нейронов разных уровней бочонковой колонки С1 от амплитуды сгибания вибриссы.
На рис.3 показан характер изменения средних размеров В-зоны РП нейронов трех уровней бочонковой колонки С3 от амплитуды сгибания соответствующей вибриссы. Следует отметить, что размер РП у нейронов слоя 4 существенно уменьшался только при амплитуде 22 мкм, тогда как у нейронов других уровней - уже при более высоких амплитудах (45 и 30 мкм), что коррелировало с их более высокими амплитудными порогами. В диапазоне околопороговых амплитуд РП нейронов слоя 4 становилось равными по размеру с РП нейронов других слоев.
Размер и структура РП обнаружили также отчетливую зависимость от уровня возбуждения бочонковой колонки. В модельной серии опытов общий уровень возбудимости мозга крыс повышался посредством внутрибрюшинного введения кофеина в дозе 100 мг/кг /24/. Так, на рис.4,А представлены результаты картирования РП нейрона, зарегистрированного в опыте N 13 на глубине 1200 мкм в бочонковой колонке С3. ПСГ в части А рисунка, представляющие центральную часть РП, построены по 32 реализациям и показывают наибольшую эффективность специфического входа в данную колонку от вибриссы С3.
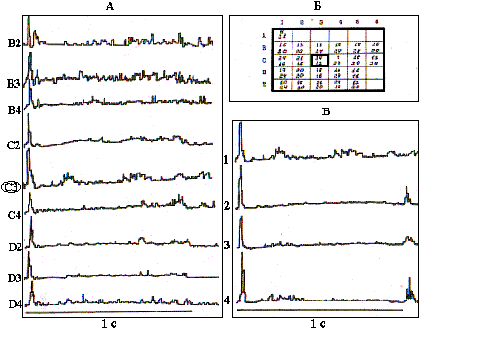
Рис. 4. Перестройка структуры В-зоны РП нейрона при повышении общего уровня возбудимости коры после введения кофеина.
А - ПСГ реакций нейрона на сгибание соответствующих вибрисс, С3 - центр РП; Б - карта РП, цифры вверху - накопление импульсов в первичную фазу, внизу - ЛП (мс) максимума ПСГ; В - формирование off- реакции на стимуляцию центра РП после введения кофеина: 1- контроль, 2- через 10 мин, 3- 20 мин, 4- 40 мин.
По всему РП в контроле данный нейрон реагирует возбуждением только на включение стимула. В части Б данного рисунка изображена полная карта РП этого нейрона со значениями максимумов в фазе первичного возбуждения и их ЛП, которые дают представление о более тонкой структуре РП. С учетом представленных показателей в РП данного нейрона также можно выделить горизонтальную в пределах рядов С и D ориентационную избирательность, проявляющуюся, прежде всего, в более высокочастотных первичных разрядах и, в меньшей мере, в их ЛП.
Через 10 мин после введения кофеина нейрон начинает реагировать также и на выключение стимула, приложенного в центре РП (рис.4,В). Со временем интенсивность первичного возбуждения на включение и выключение стимула уменьшается (к 20-й минуте), а затем к 40-й минуте вновь возрастает. Кроме того, постепенно восстанавливается вторичная активация, подавляемая на начальных этапах действия кофеина. Данный пример является весьма типичным для всей выборки исследованных нейронов (N=20). Нейроны бочонковых колонок, представляющих в С1 коры мозга крыс латеральные ряды вибрисс, имели во всех исследованных случаях односторонние РП, располагающиеся с существенным перекрытием по всему вибриссковому полю контралатеральной стороны мордочки крысы. Нейроны в представительстве самого медиального ряда вибрисс А, напротив, всегда имели билатеральные РП (N=20). В бочонковых колонках ряда В с равной вероятностью встречались и односторонние и двусторонние РП (N=10). Размеры билатеральных РП соответствовали односторонним. Их структура в общих чертах характеризовалась наличием более детерминированной по ответоспособности центральной части и периферии с низкой проявляемостью реакций на серию повторяющихся стимулов. Ипсилатеральная часть РП соответствовала периферии, однако, в ней также выделялась центральная вибрисса, симметричная центральной в контралатеральной части РП, которая давала более интенсивные и с большей вероятностью реализующиеся импульсные реакции. Кроме того, ипсилатеральная часть РП всегда ограничивалась лишь двумя медиальными рядами вибрисс - А и В. Структура импульсных реакций нейронов при стимуляции ипсилатеральных вибрисс также была фазной, с четкой тормозной паузой между первичным и вторичным возбуждением, свидетельствующей об участии системы ассоциативных связей в обработке ипсилатеральной специфической афферентации.
Перейти на страницу:
1 2 3 4 5 6 7