Контралатеральная часть РП нейронов в представительстве медиальных вибрисс, как правило, не включала в свой состав самые нижние вибриссы рядов D и Е. На рис. 5 представлено РП нейрона слоя 5 бочонковой колонки А2. Оно включает в свой состав 9 вибрисс в медиальных рядах А и В контралатеральной стороны и две вибриссы (А2 и В2) на ипсилатеральной стороне мордочки крысы. В табличках представлены значения максимумов ПСГ, построенных по 32 реализациям, а также их ЛП и длительность тормозной паузы (последние два показателя в мс). Видно, что реакции на отклонение центральных вибрисс в левом и правом полуполях РП характеризуются наибольшей интенсивностью первичного и структурированностью вторичного возбуждения. Показательны также значения тормозной паузы, максимальные для центральной вибриссы, а также значения ЛП реакций, которые в пределах всего ряда А не различались, но были достоверно (P<0,01) короче, чем у В-реакций в ответ на стимуляцию вибрисс ряда В одноименной стороны. Следует также отметить низкую проявляемость нейронных реакций в ответ на повторную стимуляцию вибрисс ипсилатеральной части РП, которая не превышала по выборке 60%, что предполагает кооперативный характер деятельности нейронов с билатеральными РП при восприятии тактильных стимулов.
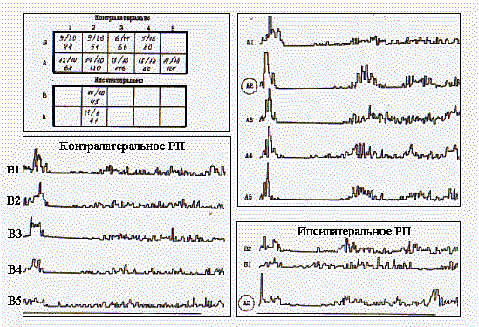
Рис.5. Билатеральное РП нейрона бочонковой колонки в представительстве медиального ряда вибрисс (объяснения в тексте).
Параметры нейронных реакций на контралатеральную и ипсилатеральную стимуляцию.
В данной серии опытов исследованы реакции нейронов бочонковых колонок представительства латеральных вибрисс в С1 коры мозга крыс на контралатеральную и ипсилатеральную стимуляцию вибрисс, а также находящейся под ними шерсти. В последнем случае в качестве топографических ориентиров при картировании РП использовались вибриссы. На рис.6,а представлены графики послойного распределения 91 нейрона, зарегистрированных в данной серии, которые реагировали первичным возбуждением на отклонение центральных в РП вибрисс контралатеральной стороны мордочки крысы (КВ-нейроны) и 54 из них, реагирующих, кроме того, на тактильную стимуляцию шерсти ипсилатеральной стороны. Мы назвали такие нейроны билатеральными (БВ-нейроны). Следует при этом отметить, что КВ-нейроны при стимуляции шерсти ипси-стороны давали такие же импульсные реакции, только более высокочастотные и коротколатентные, как и при стимуляции вибрисс контралатеральной стороны. Поэтому этот тип реагирования нейронов мы не стали выделять в качестве самостоятельного, тем более что при стимуляции шерсти не удается избежать одновременной стимуляции вибрисс (но не наоборот). Большинство нейронов представленных двух типов реагирования зарегистрировано на уровне 4 и верхней половины 5 слоев, что позволяет рассматривать их в качестве входных для афферентов вентро-базального комплекса.
На рис.6,б дано распределение по слоям коры мозга средних ЛП В-реакций, контралатерально и билатерально активируемых нейронов. При общем более низком уровне значений ЛП В-реакций нейронов первичного типа (среднее значение для всей выборки 10,3_+0,3 мс) их минимальные значения приурочены к нижней половине слоя 4 и верхней части слоя 5. Билатерально реагирующие нейроны со средним значением ЛП 18,1+0,8 мс реагируют с наименьшей латентностью в поверхностной части слоя 4 и еще меньшей латентностью в слое 6. Различие средних значений ЛП двух выборок нейронов достоверно ( Р<0,001) по критериям Стьюдента и Фишера.
Анализ статистического распределения ЛП В-реакций нейронов обеих групп, который состоял в построении вариационных рядов, их выравнивании по нормальному закону и определении достоверности различия эмпирических и теоретических распределений, показал следующее (рис. 6,в и г).
Перейти на страницу:
1 2 3 4 5 6 7 8